Toshiyuki Takahashi*1, Hisayoshi Nozaki*2
If you are asked to describe plants, what comes to mind? Terrestrial plants grow in abundance all around us, and we utilize them in our daily lives. Terrestrial plants are all members of a group called the green plants (Fig. 1)1). Red algae, which are widely distributed in the oceans and are important to humans as products such as asakusa nori (dried laver), form a different group, the rhodophytes (Fig. 1). These two plant groups have evolved complex, multicellular structures with shapes that are directly visible to the naked eye, and consequently they are very familiar organisms.
All of these plants have a common ancestor—in other words, the first plant—and it is interesting to speculate what form this must have taken. In fact, the plants that hold the key to unraveling this mystery are already known—they are a group known as the glaucophytes, which is one of the three major groups of the plant kingdom, along with the green plants and rhodophytes (Fig. 1)1-3). Although the prefix glauco- means "grey", the glaucophytes are in fact strikingly beautiful plants that range in color from blue-green to blue (Fig. 1). However, in contrast to the other two major plant groups, they are unicellular or colonial freshwater microalgae, and with no more than 20 or so species known, they are far from common3, 4).
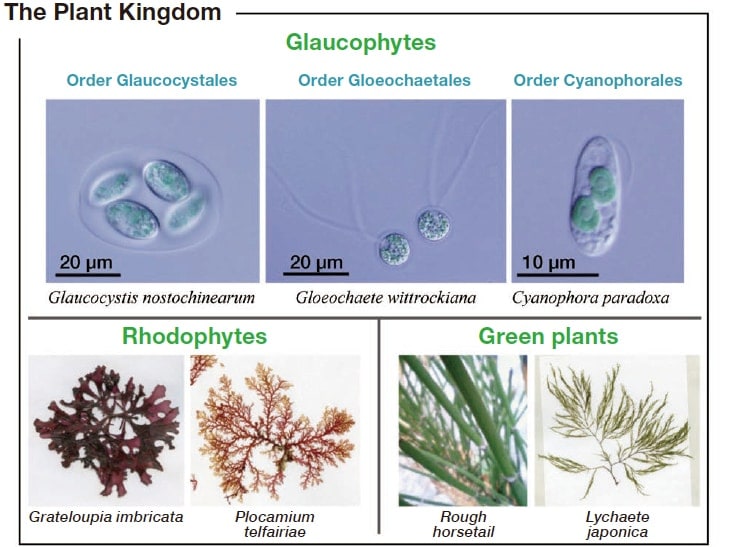
Fig. 1 The three major plant groups are green plants, rhodophytes, and glaucophytes. The glaucophytes are divided into three orders, and only a few species are known. These are freshwater microalgae, colored blue-green or blue, which can be cultured in culture medium in test tubes. From reference 1).
Plants originated 1–2 billion years ago during the Precambrian, when our animal ancestors were still swimming around as unicellular organisms1). Ancestral plants at that time had not yet evolved the capability for photosynthesis, and they are believed to have swum freely with the aid of two flagella, actively preying on their food. A heterotrophic flagellate incorporated cyanobacteria into its cell, commencing a symbiotic relationship (endosymbiosis) in which photosynthesis products of the cyanobacteria are utilized by the flagellate in return for providing the cyanobacteria with a place to live that received sunlight (Fig. 2).
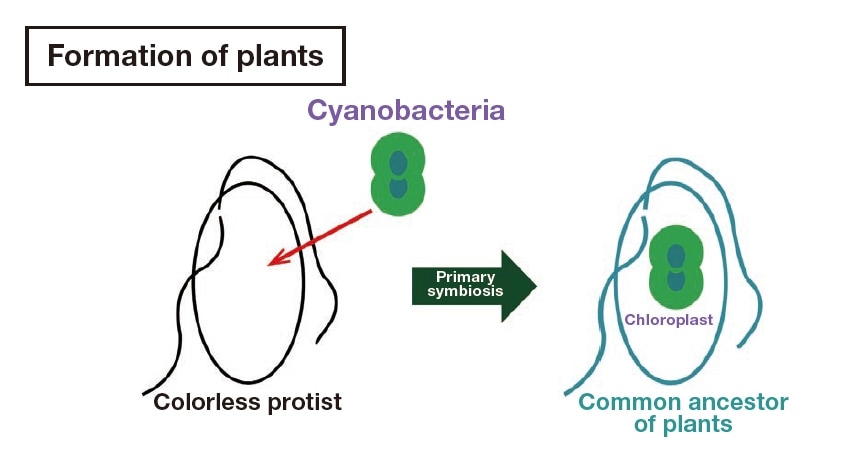
Fig. 2 Schematic diagram of the formation of plants through endosymbiosis. Between 1 and 2 billion years ago, protists that were not capable of photosynthesis incorporated photosynthetic cyanobacteria and thus became the first plant cells with chloroplasts. From reference 1).
The heterotrophs and the cyanobacteria gradually evolved to become a single, regulated organism. Thus, cyanobacteria became the chloroplast of present-day plants. This scenario of the formation of plant cells as a result of endosymbiosis—the endosymbiotic theory—is now widely accepted5), but virtually nothing is known of the lineage of the heterotrophic flagellate that incorporated the cyanobacteria. However, the chromatophore pigment composition and form that characterize present-day glaucophytes are extremely similar to those of cyanobacteria, and among the glaucophytes, the single-celled, free-swimming genus Cyanophora (Fig. 1) is considered to be a living fossil that still retains the morphology of the original plants. Cyanophora has been used in a wide variety of research, including full-genome sequencing6). Recent work has suggested that Cyanophora, which has a unique cell coat structure, retains the morphology of the first plants, possibly even the morphology of the actual flagellate that incorporated the cyanobacteria7, 8). While there have been attempts to directly capture the morphology of Cyanophora using a scanning electron microscope (SEM)9), it has not been possible to show true shape using a conventional general-use SEM.
In recent years it has become possible to culture a variety of different microalgae, such that it is no longer necessary to collect samples for observation. If a single glaucophyte individual is seeded into a test tube containing culture medium and kept in bright conditions, a culture strain comprising clones of the original cell is obtained. This strain can be maintained by sequentially seeding cells into new medium to create subcultures. Clone culture strains created in this manner are made up of genetically homogenous individuals, and such cultures are widely used in experiments as they have far greater reproducibility than samples taken directly from natural environments. Glaucophytes are uncommon and there are limited numbers of culture strains, so in order to carry out observations of cell micromorphology at the clone level, we ordered stock cultures from stock culture institutions around the world and we also established new cultures with samples collected from the natural environments3, 10).
The cells of terrestrial plants are comprised of protoplasts enclosed by a cell membrane, which in turn is enclosed by a cell wall. Microalgae, on the other hand, do not always have a cell wall, and in some cases have structures other than a cell membrane on the surface of the protoplast that are distinct from a cell wall (cell coat structures).
Glaucophytes of genus Cyanophora lack a cell wall but are known to have a unique cell coat in which the cell membrane is lined by plate vesicles, which are flat vesicles each containing a plate9). However, the fine-grained structures of the surface of this cell coat were not previously known.
We first used a general-use SEM to observe Cyanophora paradoxa cells that had been single-fixed with osmium oxide10). C. paradoxa is the most widely used species in the genus, and it has come to be used as a model of glaucophytes (Fig. 1). As in a prior study9), we were unable to observe the structure of the cell coat on the surface of the cell with an accelerating voltage of 15 kV. At low accelerating voltages, however, it was possible to observe the cell surface structures because the electron beam did not penetrate the cell surface. Consequently, the presence of a fine-grained structure on the surface of the cell coat was shown (Fig. 3)10).
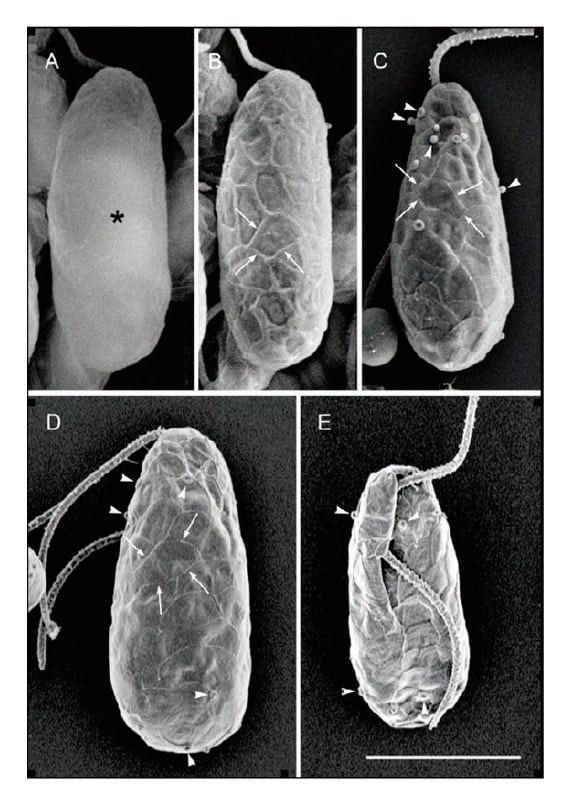
Fig. 3 SEM images of the glaucophyte Cyanophora paradoxa. Comparison of SEM techniques. Bar = 5 μm.
However, general-use SEM did not have sufficient resolution to show detailed patterns of the cell coat of Cyanophora, and we therefore carried out observations using an ultra-high resolution field emission (FE)-SEM (model SU8020) at the model room of Hitachi High-Tech Corporation. Recent ultra-high resolution FE-SEMs allow lower landing voltage than the applied voltages of the field emission gun, and we hoped that an FE-SEM capable of low accelerating voltages and ultra-high resolution observation would reveal the details of the fine-grained structure of the cell coat surface. The resulting images showed a pattern bordered by ridges covering the whole cell surface of C.paradoxa (Fig. 3)10). We had never imagined that the whole surface of the cell membrane of Cyanophora would be covered with this minutely detailed pattern, so the observation came as a great surprise. We carried out transmission electron microscope (TEM) observation of ultra-thin sections in order to elucidate the structure of the ridges, and we found that they are formed by upward bending at the border between the plate vesicles that line the cell membrane10). Cyanophora cells lack a cell wall, and it therefore appears that the flat vesicles containing firm plates line the inside of the cell membrane with no gaps, providing support to the cell. The plate vesicles overlap one another, causing the minute structure of ridges and patterns on the surface of the cellular membrane. Ultra-high resolution FE-SEM thus captured this detailed micro-pattern produced by flat vesicles distributed across the entire inside of the cell membrane. From our subsequent research, we conjecture that a Cyanophora-like flagellate having a cell coat structure of leafletlike flat vesicles closely lining the inside of the cell membrane was the common ancestor of the glaucophytes and, by extension, all plants8). Our FE-SEM observation of C. paradoxa gives a glimpse at the level of fine-grained structure of the first plant from 1–2 billion years ago.
It should be remembered that the genus Cyanophora does not only comprise the single species of C. paradoxa. If we do not grasp the species diversity of this genus, we will be unable to appreciate the shared traits that extend back to the morphology of their common ancestor, the first plant. The glaucophytes have evolved at the same time as green plants and rhodophytes, and we have conducted taxonomic research into glaucophyte species with the expectation that they will also exhibit some degree of morphological diversity2). Three species of Cyanophora have traditionally been described, but no studies have looked at the micromorphological diversity of the many clone strains3). We therefore commenced comparative morphological observations with a focus on micromorphology that included the cell coat structure found in C. paradoxa. Just as species classifications have been drawn up for macroscopic plants, we hoped that using the latest electron microscope equipment would enable us to establish a species classification by morphology even for the minute glaucophytes2).
We carried out our morphological comparison using low accelerating voltage FE-SEM and ultra-thin section TEM on a total of 8 culture strains of glaucophytes, including a new strain that we established independently (Fig. 4).
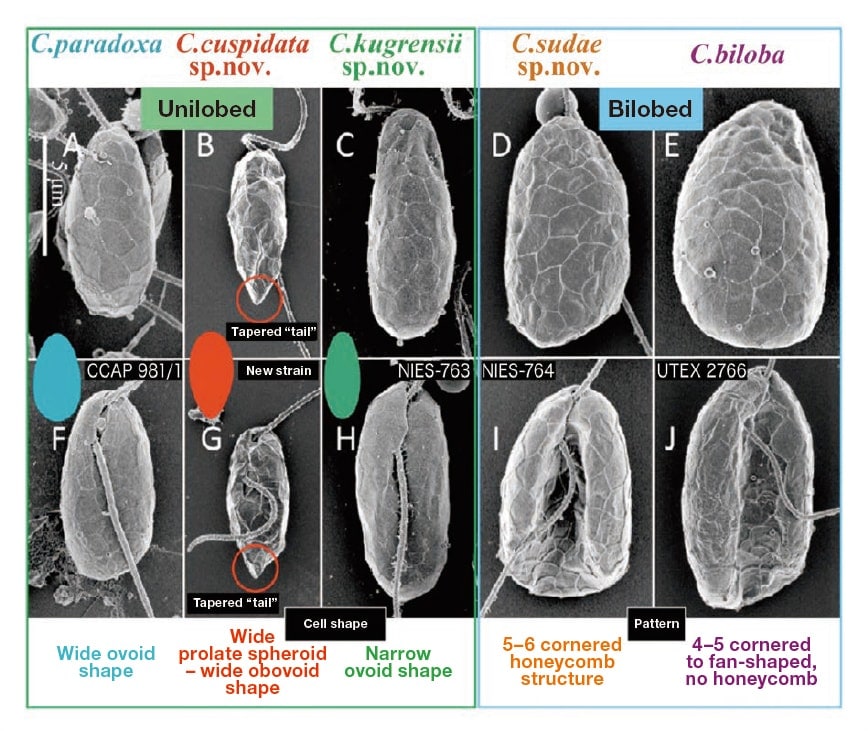
Fig. 4 Low accelerating voltage field emission scanning electron microscopy (FE-SEM) photographs of 5 species of genus Cyanophora. Bar = 5 μm. (A, F) Cyanophora paradoxa. (B, G) New species C. cuspidata. A highly tapered tail sometimes forms at the tip of the cell. (C, H) New species C. kugrensii. (D, I) New species C. sudae. The pattern has a 5–6 cornered honeycomb structure. (E, J) C. biloba. The pattern ranges from 4–5 cornered to fan-shaped. Reprinted with permission of John Wiley and Sons (License Number: 3790920895102) from Reference 3).
The results show that the previously mentioned fine-grained structure that was established for C. paradoxa, with plate vesicles that line the cell membrane overlapping each other and completely covering the cell, is a common trait shared across the genus3). Simultaneously, the genus can be broadly divided into bilobed species, in which the cell is divided into left and right lobes by a deep groove running down the center of the cell when viewed ventrally, and unilobed species, in which no such differentiation into lobes is visible (Fig. 4).
Comparison of the pattern enclosed by ridges on the surface of the cell from the dorsal side of the cell in the bilobed group shows that while C. biloba has a pattern ranging from 4–5 cornered to fan-shaped, the new species C. sudae has a pattern of 5–6 cornered honeycomb shapes, and the two species can be distinguished based of this difference in surface pattern (Fig. 4)3). The cell membrane was detached and observations made using freeze fracture TEM showed that the pattern observed by low accelerating voltage FE-SEM reflected the morphology of the interior plate vesicles (Fig. 5).
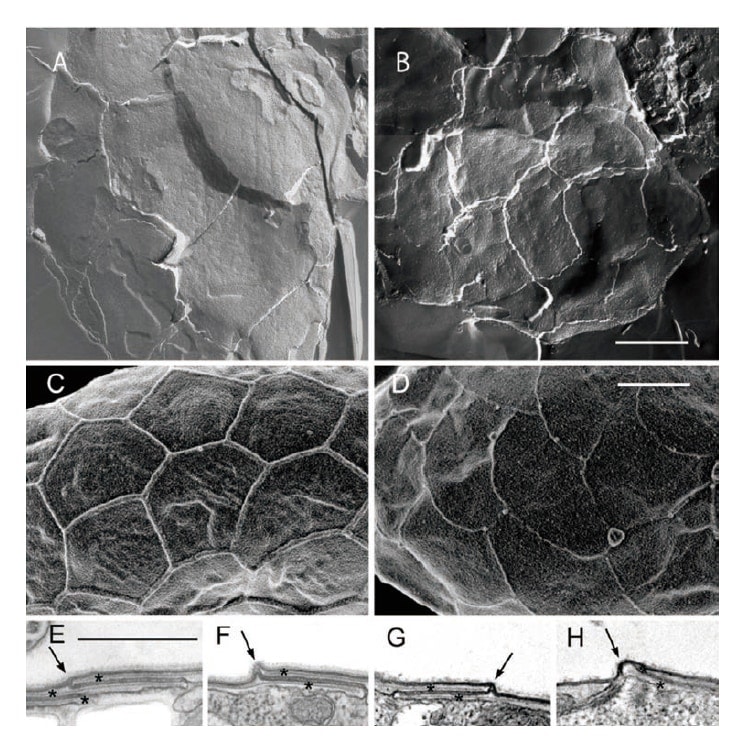
Fig. 5 Images by freeze fracture transmission electron microscopy (TEM) (A, B), field emission scanning electron microscope (FE-SEM) (C, D), and ultra-thin section TEM (E-H). The pattern covering the whole of the cell surface is bordered by ridges, which are formed by upward bending at the boundary between plate vesicles such that the pattern reflects the morphology of the internal plates. Bar = 1 μm. (A, C, E, F) New species Cyanophora sudae. (B, D, G, H) C. biloba. Reprinted with permission of John Wiley and Sons (License Number: 3790920895102) from Reference 3).
In the unilobar group, new species C. cuspidata was differentiated by most specimens having an extremely tapered "tail" at the tip of the cell (Fig. 4)3). This tail was clearly shown for the first time to be extremely sharp and to be a characteristic of this species through the use of low accelerating voltage FE-SEM. In addition, the cell shape was measurable in SEM photographs that clearly showed the contours of the cell, allowing identification of statistically significant differences in morphology between species. In C. cuspidata, the front end of the cell was more swollen with a (top heavy) wide prolate spheroid to wide obovoid shape. C. paradoxa had a stout, egg-shaped wide ovoid shape, and the new species C. kugrensii had a narrow ovoid shape, like a long, thin egg (Fig. 4).
The taxonomic study presented here3) identified three new species (Fig. 4), establishing that there are twice as many Cyanophora species as previously thought. This suggests that the glaucophytes, which have always been considered to be a very small taxonomic group, may include a considerable number of undescribed species that cannot be distinguished by optical microscopy alone. Morphological comparison using low accelerating voltage FE-SEM is effective for discovering taxonomic characters that allow species to be distinguished. This technique has the advantage that the whole cell can be observed, which was used to clarify identifying characteristics at the level of the finegrained structure. An FE-SEM, which allows observation at higher magnification, revealed interspecies differences at nanoscale in the roughness of the cell membrane surface. However, under the present conditions, these were not adopted as taxonomic characters2). In the near future, it is likely that the number of culture strains of this genus that can be used will increase and additional new species will be distinguished based on nanoscale taxonomic characters.
As we have outlined thus far, morphological comparison using low accelerating voltage FE-SEM clarified taxonomic characters at the level of fine-grained structure, demonstrating this to be a powerful tool for distinguishing microalgae species. This method also allowed us to "see" the shape of the first plants from 1–2 billion years ago, something that we cannot directly observe in the present day. If more glaucophytes and eukaryotes can be classified to species based on the level of fine-grained structure, it may become possible to conjecture their past morphology and evolution process2, 8). We are currently working on a species-level taxonomic study of genus Glaucocystis, a different type of glaucophyte (Fig. 1)1).
Unlike Cyanophora, Glaucocystis possesses a thick cell wall8) such that the cell coat cannot be directly observed by low accelerating voltage FE-SEM. For this reason, we carried out observations of the cell coat structure at the 3-dimensional fine-grained structure level by means of tomography using an ultra-high-voltage electron microscope (H-3000). We have also made observations of the fine-grained structure of the cell wall surface using ultra-high resolution FE-SEM (SU8220). To date, comparative morphological observation by means of ultra-high-voltage electron microscopy has not been used in the classification of microalgae, but as this technique allows direct observation of the 3-dimensional structure of protoplasts within the cell wall, it is likely to be extremely helpful in the future for classification of microorganisms based on analysis of the diversity of fine-grained cell structure.
In 2015, an atomic-resolution holography electron microscope, which had long been the earnest wish of the late Dr. Akira Tonomura, was completed11). In the field of biological samples as well, detailed ultra-high resolution observations of whole surface structure by FE-SEM and scanning probe microscopes (SPM) have become possible as a result of recent technological innovations, and 3-dimensional information such as roughness on the cell surface can now be readily obtained. At the same time, the age in which we can obtain 3-dimensional information of the inside of the cell in its entirety by using ultra-high-voltage electron microscopes or focused ion beam (FIB)-SEM has now arrived. These next-generation ultrafine morphological observations are expected to become the mainstream for future microorganism classification, superseding the fragmentary information that conventional electron microscopes were only able to provide. Through the merger of taxonomy and next-generation micromorphology, ultrafine morphology at the 3-dimensional level of present-day and ancestral organisms will likely be clarified. This field has high expectations of ever greater technological innovations in electron microscopy.
Acknowledgements
We are grateful to Dr. Mayuko Sato and Dr. Kiminori Toyooka (Riken) for their painstaking instruction of low accelerating voltage SEM techniques and fixation methods as joint research. We carried out joint research into freeze fracture TEM observation at Kochi University under the kind guidance of Mai Kawamura and Professor Kazuo Okuda. FE-SEM was carried out using the SU8020 at the model room of Hitachi High-Tech Corporation, and we were assisted in making observations by Yukari Dan and Shunya Watanabe. Dr. Kaoru Kawafune (Tokyo Institute of Technology) helped with the algae samples, and Dr. Rei Matsuzaki (The University of Tokyo) went to very great lengths to help in many different ways, from handling the microalgae to molecular experiments.
References
About the author
*1 Toshiyuki Takahashi
Completed doctorate course at Department of Biological Sciences, Graduate School of Science, The University of Tokyo. Doctorate (Sciences)
*2 Hisayoshi Nozaki
Associate Professor at Department of Biological Sciences, Graduate School of Science, The University of Tokyo. Doctorate (Sciences)
Original paper written in Japanese “The Ancient Plant Cell Now " Visible" – Native Ultrastructural Feature and Diversity of Glaucophytes Unveiled by Ultra-High Resolution FE-SEM” has been translated to this English version by Hitachi-High Tech Corporation.
See more
