MirrorCLEM: Seamless System for Correlative Light and Electron Microscopy

国立研究開発法人 理化学研究所
環境資源科学研究センター(CSRS)
技術基盤部門 質量分析・顕微鏡解析ユニット
豊岡 公徳 博士(理学)
生体内のタンパク質等の機能分子の働きを正確に知るためには、それらの細胞内局在パターンを明らかにする必要がある。例えば、特定のタンパク質の細胞内局在やその動態は、緑色蛍光タンパク質(GFP)等を用いた蛍光イメージングの発展により、近年は容易に観察できるようになった。しかし、光学顕微鏡(光顕)の解像度以上の微細な細胞内構造情報が必要なケースでは、電子顕微鏡(電顕)を用いた観察が必要となるが、電顕では蛍光イメージングで用いる可視光領域を検出することができない。そのため筆者らはこれまで、電顕レベルで蛍光標識タンパク質の細胞内局在を解析する場合、蛍光タグに対する抗体を用いた免疫電顕法による局在推定を行ってきた1)。しかし免疫電顕法は、抗体・抗原の性質によっては使用できない場合も多く、高度な技術と経験を必要とする手順を含む。さらに金粒子が局在する構造体が、本当に蛍光を発する構造体と同一であると断定することは難しく、同一であると“推定”することしかできないのが実状である。この“推定” から“ 特定” にステップアップするためには、GFP などの蛍光を観察したのと全く同じ試料を電顕観察する必要がある。それを可能とする技術が、光- 電子相関顕微鏡法(Correlative light and electron microscopy: CLEM)法である。
CLEM 法は、同一箇所を光顕と電顕を用いて観察し、両顕微鏡により得られた像を重ね合わせることで相関を得る解析法である。光顕では、蛍光試薬や色素による組織染色など分子の局在・分布を広範囲で可視化できる。一方、電顕では、光顕では見えない超微形態を可視化できる。光顕と電顕を組み合わせるCLEM 法では、特定視野の分子局在と超微形態情報の両面で説得力のある実証性の高いデータを提示できる。生物試料では、蛍光免疫染色法や組織化学染色法などと透過電子顕微鏡(TEM)や走査電子顕微鏡(SEM)と組み合わせたCLEM、材料系試料では、反射顕微鏡像とSEM を組み合わせたCLEM がある。これまで多くのCLEM 法が報告され、顕微鏡メーカーから発表されている。筆者の理想的なCLEMは、メーカー問わず全ての光学顕微鏡像と電顕像が相関可能な、迅速で正確なCLEMシステムである。本稿では、(株)日立ハイテクと開発したSEMとTEM に対応したCLEMシステムMirrorCLEMとその応用について報告する。
筆者が初めてCLEM 法にチャレンジした時は、電顕観察には電界放出形走査電顕(Field emission SEM: FE-SEM)を用いたが、光顕で得られている特定視野の蛍光像と同一箇所をFE-SEM で探すことは非常に難しかった。その際は、切片試料の形やカバーガラスに印刷された英数字付きグリッドを参照して同一箇所を探し出し、画像処理ソフトを用いて蛍光像とFE-SEM 像を重ね合わせて辛うじて相関像を得たが、撮像後も同じ倍率・同じ傾きで重ね合わせるのに時間を要した2)。そこで筆者は、(株)日立ハイテクと共同研究開発を行い、迅速で簡便なCLEMシステム“MirrorCLEM” を開発した3)。本システムは、図1 に示すように、①光学顕微鏡を用いて樹脂切片などの試料を低倍率からターゲット構造が明瞭に観察される高倍率まで撮像した後、② MirrorCLEMソフトウェアによって1枚の光顕マップを作成、③観察位置情報をFE-SEM またはTEM のステージ位置情報と同期させたうえ、④ MirrorCLEMソフトウェアからステージの外部制御を行うことで、電顕による同一箇所の観察・撮像・重ね合わせができる。また、光学顕微鏡とFE-SEM のオーバーレイ画像のリアルタイム表示や追従も可能である。MirrorCLEM を使えば、撮影した光顕と電顕のステージが鏡を見ているかのようにシームレスに観察・撮像することができ、効率よく相関像を得ることができる。次に、SEMと組み合わせたMirrorCLEM の応用と、その後に、TEMと組み合わせたMirrorCLEM の応用について紹介する。
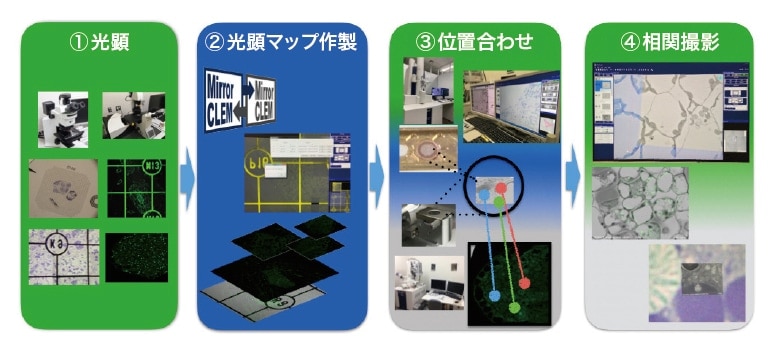
図1 光-電子相関顕微鏡システム「MirrorCLEM」のワークフロー図
近年のSEM は電子銃や検出器など性能が上がり、生物試料を含む樹脂包埋切片をスライドガラスやウェーハーなど平板上に載せ、低加速電圧ビームと反射電子検出器を用いて観察する“ 切片SEM 法” により、TEMと遜色ない細胞・細胞内小器官の切片電顕像を撮影できるようになった4,5)。これにより、TEM で用いる超薄切片でなく200 nm 以上の厚い樹脂切片を用いて、広域に渡り容易に撮像することができる。FE-SEM は、電界放出形電子銃を用いることにより、高分解能で数十万倍の倍率での観察が可能であり、筆者が使っているHitachi SU8220は、20-30 kV の高加速電圧で高分解能像を得られるだけでなく、0.1-1 kVといった極低加速電圧条件下でも大きなプローブ電流が得られ、明るくS/N に優れた画像観察が可能な反射電子検出器、YAG 結晶を用いた高感度の反射電子検出器(YAG-BSE)6)など搭載されている。1 μm 以上の厚い切片でもスライドガラスに載せ、電子染色後、オスミウム蒸着し、YAG-BSE 検出器を用いることで、帯電なくスムーズな観察が可能である7)。
SEM-MirrorCLEM(図2A)は、光顕で一般的に使われるスライドガラスまたはカバーガラス専用の試料台(図2B)と、MirrorCLEMソフトウェアからなり、FE-SEMステージ上の切片と蛍光写真など光顕像の位置を3点アライメント法により容易に同期することができ、同一箇所の微細構造を撮影できる。さらに、MirrorCLEM から光顕のX-Y 自動ステージを制御し、低倍率から高倍率の光顕像を取り込むことで、光顕マップとFE-SEM のステージとの位置合わせを簡便に行うことも可能である(図2C)。
FE-SEM を用いたMirrorCLEM 観察は、先に切片などの試料を光顕で色情報や低倍率像を取得したのち、同一試料、同一箇所をFE-SEM で撮像して、光顕像と重ね合わせ、相関像を得る。紙や基盤などの材料系であれば、先に光顕で撮影して、金属コーティング後にFE-SEM に入れ、アライメントを取るだけで簡単に相関像が得られる。生物試料の場合は、固定・樹脂包埋してウルトラミクロトーム等で作製した樹脂包埋切片をスライドガラスに載せ、色素染色または蛍光を光顕で検出したのちに、電子染色およびオスミウム蒸着し、FE-SEM の反射検出器で撮影することで観察することで、同一切片をSEM 観察できる。ここでは生物試料の種類・目的に応じて使い分けている[1] 〜[3] の3つの方法を紹介する。
オスミウム固定試料を用いた形態観察用樹脂ブロックから500 nm-1 μm の準超薄切片を作製し、スライドガラスに貼り付ける。TB 染色し、正立顕微鏡等で低倍像と興味ある場所(Focus of view: FOV)の高倍像を撮像する。その後、酢酸ウラニル溶液とクエン酸鉛溶液で電子染色し、オスミウムコーティングを行う。MirrorCLEM に低倍〜高倍率の光顕像を読み込み、光顕マップを作成する。スライドガラス用SEM 試料台にスライドガラスをセットし、MirrorCLEM で切片と光顕マップの3点アライメントを行う。これにより、光顕マップを移動するとステージも同じ場所に移動し、光顕マップを拡大するとSEM も同様に同じ倍率に拡大され、撮像すると自動に光顕マップ上にSEM 像を重ねて保存される。例えば、アズキの発芽子葉のTB 染色切片とそのFE-SEM 像を撮像することで、光顕で観察位置を把握しながら光顕では見えない細胞内小器官を撮像することができた(図2-[1])。
動物の接着細胞はグリッド付カバーガラス(プリント面)に貼り付け、グルタルホルムアルデヒド固定後、共焦点レーザー顕微鏡等で蛍光と走査像のグリッドの英数字を含む低倍率像とFOV を含む高倍率像を撮影する。その後、オスミウムで固定、脱水、エポン樹脂に倒立包埋する。カバーガラスを樹脂から剥がすと、英数字と枠の凹凸が残るため、その凹凸を指標に光顕で撮像した箇所を残してトリミングし、超薄切を行う。その切片をスライドガラスに載せ、電子染色とオスミウムコーティング後、[1] と同様にMirrorCLEM で細胞やグリッド枠を指標にアライメントを行い、相関像を得る(図2-[2])。この方法を用いて、マウス皮膚初代培養細胞を用いて赤色蛍光タンパク質でラベルしたケラチンのCLEM 解析に成功した8)。
スライドガラスに接着できない組織や浮遊細胞のGFP など蛍光タンパク質の局在を観察する場合は、その蛍光を残して樹脂包埋と超薄切を行う。その切片をグリッド付カバーガラスに載せて蛍光観察し、その同一箇所をMirrorCLEM 搭載FE-SEM で観察する(図2-[3])。例えばGFPラベルしたERボディ(小胞体由来の顆粒)またはペルオキシソームを含む形質転換体シロイヌナズナを用いてCLEM を試みた。芽生えをアルデヒド固定し, 脱水後にアクリル系樹脂であるLRWhite 樹脂に包埋し、準超薄切片(厚さ1 μm)を作製し、グリッド付カバーガラスに切片を載せ、共焦点レーザー顕微鏡で蛍光像を撮影した。電子染色後、同じ切片のGFP 蛍光が検出された箇所をMirrorCLEM 搭載FE-SEM で撮影した。その結果、蛍光ドットは、紡錘形のERボディまたは丸いペルオキシソームと一致する相関像を得た9)。

図2 SEM-MirrorCLEMおよびその試料調製法
(A)当施設のMirrorCLEMシステム。MirrorCLEM搭載SU 8220および自動ステージを搭載した倒立顕微鏡。
(B)スライドガラスおよびカバーガラス用FE-SEM試料台。
(C)MirrorCLEMによる光顕自動ステージ。
[1]-[3] 生物試料の試料・目的による相関解析法のワークフロー。
透過電顕HT7800など最新のTEM を制御できるMirrorCLEM(図3A)は、光顕で撮影した像を読み込み、3点アライメントを取る(図3B)ことで光顕像上の興味ある場所をTEM で探す、あるいは電顕で光顕像のどこを観ているか確認しながら、光顕像に同一箇所を撮像し、保存することができる。相関像を得られるだけでなく、光顕と電顕像をリンクさせて保存できるので非常に便利である。ここでは、[1] 準超薄切片と超薄切片とのCLEM、[2] TEMグリッド上の蛍光微粒子のCLEM、[3] TEMグリッド上に載せた超薄切片蛍光タンパク質蛍光像とのCLEM、の3つの具体例を紹介する。
連続した切片同士を用いて相関を取るTEM-CLEM 法、つまりスライドガラス上の準超薄切片像とその直後のTEMグリッドに載せた超薄切片を用いて相関像を得る方法である。例えば、高圧凍結/ 凍結置換したシロイヌナズナ鞘の細胞内小器官を観察する場合、まず包埋した樹脂ブロックの準超薄切片を作製し、最後の1枚をTB 染色し、正立顕微鏡で低倍率とFOV の高倍率写真を撮影する。その後、FOV を含む部分を残してカミソリでトリミングし、80-100 nm 厚の超薄切片を数枚作製する。そしてTEMグリットに載せる。電子染色を行い、MirrorCLEM 搭載HT7800にセットし、アライメントを行うことで容易に同じ場所を撮影する(図3-[1])。TB 像にTEM 像を重ね合わせて表示することで、鞘のどの箇所を拡大し撮影したゴルジ体やミトコンドリアか特定することができた(図3C)。
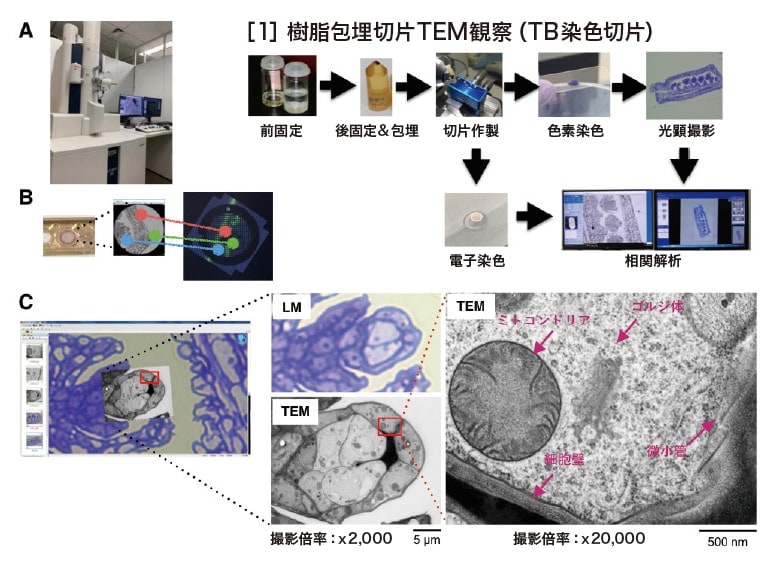
図3 TEM-MirrorCLEMおよびその試料調製法
(A)MirrorCLEMを搭載したHT7800。
(B)グリッドと光顕像との3点アライメントのイメージ図。
(C)高圧凍結/凍結置換したシロイヌナズナ鞘のTB染色像とMirrorCLEM像。
[1] 樹脂包埋切片TEM観察(染色切片)のワークフロー。
蛍光を放つ微粒子などはTEMグリッドに載せ、蛍光撮影したのちにネガティブ染色を行いTEM-MirrorCLEM で観察を行う。例えば、蛍光ビーズの場合、浸水化処理した膜張TEMグリッドに蛍光ビーズを載せ、乾燥させる。そのグリッドをスライドガラスとカバーガラスに挟み、共焦点レーザー顕微鏡で蛍光および走査像を撮像する。酢酸ウラニル水溶液液でネガティブ染色後、TEM-MirrorCLEM を用いてアライメント後、CLEM 撮影する(図4-[2])。緑色蛍光を放つビーズとそうでないものをTEM で識別することができた。
GFP 蛍光を残すようにマイルドに固定包埋した樹脂ブロックを作製9)し、100-200 nm の超薄切片を作製し英数字グリット付TEMグリッドに載せる。蛍光を復帰させるために退色防止剤で満たしたカバーガラスボトム35 mmディッシュとカバーガラスに挟み、高感度検出器を搭載した共焦点レーザー顕微鏡で蛍光像と走査像を撮像する9)。その後、電子染色し、TEM-MirrorCLEM でCLEM 撮影を行う(図4-[3])。例えば、高感度共焦点レーザー顕微鏡像で英数字グリット付TEMグリッドに載せたシロイヌナズナ発芽子葉中のGFPラベルしたペルオキシソーム樹脂包埋超薄切片の同一切片・箇所をMirrorCLEM で探し(図4A)、TEM 像を撮像し蛍光像と重ねる(図4B)。どの構造がペルオキシソームか区別することができた。
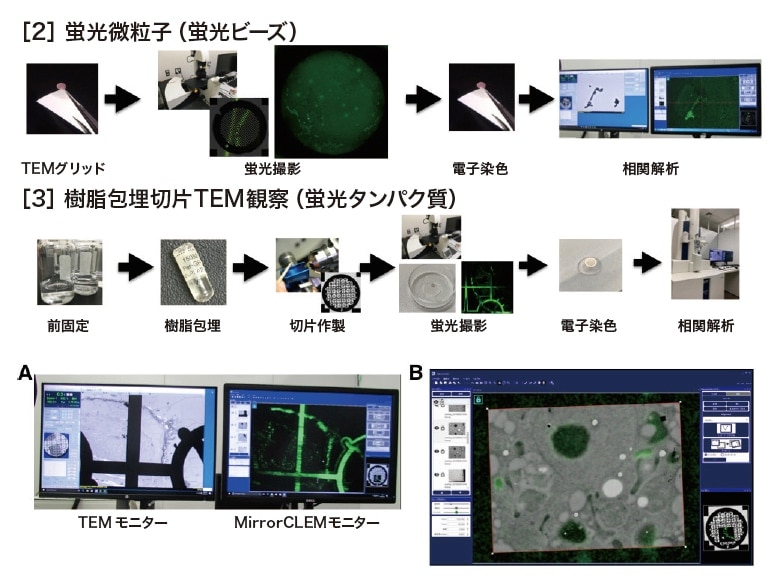
図4 TEM-MirrorCLEMの試料調製法
[2] TEMグリッド上の蛍光微粒子の相関像撮影のワークフロー。
[3] 蛍光タンパク質GFPの蛍光とTEMでCLEMする場合のワークフロー。
(A)同一箇所を観察中のTEMモニター画面とMirrorCLEMモニター画面。
(B)GFPラベルしたペルオキシソームの共焦点レーザー顕微鏡像と同一箇所を撮像したTEM像の重ね合わせたMirrorCLEM画面。
MirrorCLEM は、蛍光タンパク質や色素が局在する微細構造を簡便に特定でき、多種多様な像と相関が得られるシームレスな電顕システムである。近年では電顕と光顕で同じ範囲、近い解像度で観察できるようになり、共焦点レーザー顕微鏡やFE-SEM の検出器の向上により、今まで得られなかったシグナルを高感度で検出できるようになった。今後は、共焦点レーザー顕微鏡および連続切片SEM 法による3次元解析法とMirrorCLEM を組み合わせて解析することで、標的タンパク質の3次元的な局在様式が超微形態レベルで明らかにできると期待される。さらにMirrorCLEMと高圧凍結・クライオ電顕など他の様々な新技術を組み合わせて多角的な視点からの超微形態観察を行うことで、生体分子の分布や動態が極めて緻密に明らかになり、生命現象の理解に大きく貢献できると期待される。
参考文献
謝辞
本システムの開発にあたり、(株)日立ハイテクの皆様、(株)アストロンの皆様、理研CSRS の成川苗子氏、浜村有希博士、若崎眞由美氏、佐藤繭子博士に多大なるご協力を頂きました。また、基礎生物学研究所の真野昌二博士にGFP 発現シロイヌナズナを分与頂きました。理研IMS の松井毅博士、臼井景子博士、井関八郎氏にGFP 発現マウス皮膚初代培養細胞を提供頂きました。この場を借りてお礼申し上げます。本研究は科学研究費 新学術領域研究(17H06475)および基盤C(17K07477)のサポートにより行われました。
さらに表示






